
Generic selectors Exact matches only Search in title Search in content Post Type Selectors Learn More

Roblox: All Weak Piece codes and how to use them (Updated December 2022)

With countless beloved anime series adapted as an experience on Roblox , there’s almost always something for everyone. If you love the anime One Piece , the adaptation of the number 1 best-selling manga in history, then you’re in for a treat. There are a couple of games on Roblox that are based on the beloved series.
One game that will definitely interest you is Weak Piece by Shonen Production. Based on One Piece , you will eventually get your hands on devil fruits such as Goro , Magma , Mochi on your way as captain of the seven seas. You will also get to use weapons such as the Excalibur !
However, your journey won’t always be easy, and you may need to use some codes to give you some goodies to win your battles. If that’s the case, you’ve come to the right place. This article is dedicated to gathering codes that you can use in Weak Piece .
READ MORE: Roblox: All Southwest Florida Codes and How to Use Them
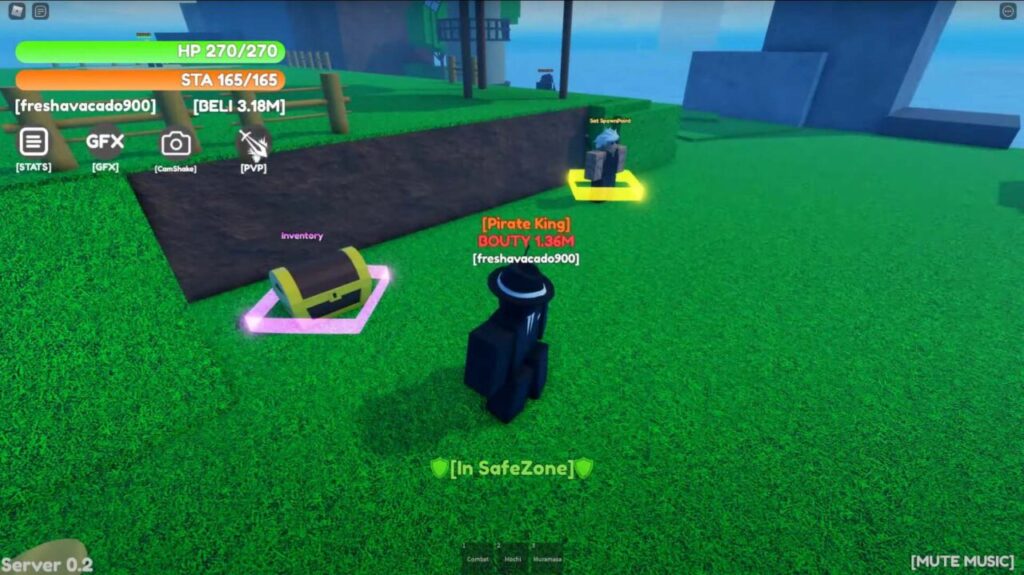
List of All Weak Piece Codes
Below is a list of codes that will definitely help you in your journey across the seven seas. Make sure you enter the codes as they are because they can be case-sensitive.
Moreover, redeem these codes as fast as you can . You never know when they will expire.
ACTIVE CODES
These Weak Piece codes are still valid. If one doesn’t work, we will make sure to update this article.
- ( There are currently no active codes. )
READ MORE: Roblox: All Combat Warriors Codes and How to Use Them
EXPIRED CODES
The codes below are no longer valid. We will also make sure to update the article once some of the current active codes expire.
- ( There are currently no expired codes. )
How to Use Codes
As there are still no available codes, we still cannot provide you with the actual steps to use your Weak Piece codes at the moment. We will make sure to update this article once codes are available. Stay tuned!
Roblox is an online game platform and game creation system. Released in 2006, it allows users to create games for others to play and/or play games created by others.

All Rights Reserved. No part of this site or its content may be reproduced without the permission of the copyright holder.
Genshin Impact
Grand theft auto, click media group, battle pass, advertise with us | write for us | privacy policy | opencritic | brands | contact us, © 2024 click media group | all rights reserved | proudly developed by darkdev. design.

Generic selectors Exact matches only Search in title Search in content Post Type Selectors
The Wandering Key: A Closer Look
Presiding over the east end of the Music Concourse, the Francis Scott Key Monument is one of the largest monuments found in Golden Gate Park. However, where you see it today is not its original location and it had a bit of a troubled history before it found its way there.
The Key Monument was a gift to San Francisco from James Lick. One of the richest men in California, Lick died on October 1, 1876. In his trust, he set aside $60,000 for a monument to Francis Scott Key in Golden Gate Park to honor the composer of the Star Spangled Banner. It took over eight years before William Wetmore Story was awarded a contract to build the monument in February 1885. Story created the monument in Italy over the next two years before having it shipped to America in numbered pieces.

Story’s travertine monument featured a sitting Key on a large base that has verses from the national anthem on it. Above the four archway views of Key stands a flag-holding Columbia with eagles on each corner. The monument stood 51 feet tall. There was discussion of several sites for the monument–atop Strawberry Hill and at the Haight Street entrance for two–before the City settled on a spot near where the new music stand was to be built.

Both the Key Monument and the new music stand were unveiled in a big dedication on July 4, 1888. Mayor Edward B. Pond accepted the monument from the Lick Trust at the ceremony attended by more than 10,000 people. A military band played, the National Guard gave a 100-gun salute, and the crowd sang the Star-Spangled banner naturally. In his remarks, Pond declared: ” It has fallen to the lot of San Francisco, through the generosity of one of its citizens, to be the first to do honor to [Francis Scott Key’s] name by the erection of a monument to his memory. 1 ”
Despite the dedication of the Key Monument, Story had not been fully paid for his work. The Lick Trustees had expected several bronze bas reliefs on the pedestal, one depicting the battle of Fort McHenry (the inspiration for Key writing the Star Spangled Banner) and another depicting a choir singing the anthem. Story had put slabs with inscriptions of the anthem on the base of the monument instead. Apparently the bas reliefs were not considered works of art under U.S. law and could not be brought to San Francisco duty-free, so Story, who had complete artistic discretion on the project, changed the concept slightly. 2

However, the Key monument’s first location became temporary for the same reason that many other things changed in San Francisco. On April 18, 1906, the Key Monument, like much of San Francisco, was heavily damaged by the earthquake. Further, in the 18 years since the dedication of the Key Monument, trees had grown up around its location and it was decided that the placement was no longer sufficiently prominent. In January 1908, the Park Commissioners took a drive through the Park to pick a new spot for the Key Monument and decided on a spot on the south side of the Music Concourse.

In March 1908, the McGilvray Stone Company was hired to move the Key Monument to the new location on the Music Concourse. The restoration of the monument would continue into 1909. In 1916, the California Academy of Sciences moved into the North American Hall of Birds and Mammals by the Key Monument. The Academy expanded with the addition of the Steinhart Aquarium in 1923, the Simson African Hall in 1934, a Science Hall in 1951, and the Morrison Planetarium in 1952.
The Key Monument was moved again in 1967 to make room for further expansion of the Academy of Sciences, this time into storage at the Park’s Corporation Yard. It would sit there for 10 years. Raymond Clary, who literally wrote the book on how the Park was created, 3 fought to get the monument back on display in the Park. On the 89th anniversary of its original installation, the Key Monument was rededicated on July 4, 1977 at the east end of the Music Concourse. Five days later, on July 9, 1977, it was designated as San Francisco Landmark #96.
You can learn more about the Key Monument in the Outside Lands Podcast we did on it . It remains there today and recently received a $140,000 renovation. Earlier this year, a ferris wheel was partially constructed behind it for Golden Gate Park’s 150th anniversary, though construction was halted due to the pandemic shelter-in-place requirements. Maybe the Key Monument’s wandering days are now over, but only time will tell.
1. “Francis Soctt Key, His Monument Unveiled at the Park,” San Francisco Chronicle, July 5, 1888, p. 8.
2. “The Lick Trust. Its Affairs In A State Of Repose” San Francisco Chronicle, January 12, 1889, p. 5.
3. “The Making of Golden Gate Park, The Early Years: 1865-1906” by Raymond H. Clary (California Living Books, 1980).
How & When to use Weak key and value?
https://developer.roblox.com/articles/Metatables
I know how to use MetaTables but I don’t get the concept of _mode what is it useful for and how do I use it?
So when / why / how do you use Weak Keys/Values?
For Roblox, you never really need to.
Some people like to use instances as keys for things like:
…where coins has keys of Player instances and values of their coin amount. They then learn that when the player leaves, this doesn’t GC the object. So, they try this:
The idea is that the rest of your code is normal, but when the player leaves (and is parented to nil), it’ll clear out the key. However, that’s not the case because Weak Tables don't work with Instances .
The actual solution is just to hook on PlayerRemoving and set the data to nil there.
The point is that you never need weak tables and using them only really causes problems down the line; there’s almost always a better and more explicit solution.
So it’s basically useless then?
That is a shame.
Can you show me an Example of how to use it with something else that aren’t Instances?
I used to ask questions on a website called scripting helpers . I asked this question: Weak Tables . EgoMoose explained it really well with their answer. It helped me understand weak tables.
So let’s say you have a table in a script whose lifetime is as long as the server and whose keys are all of the hats of every character in the game, mapped to the player the hat belongs to. It might sound absurd but I’ve written it before.
I had what was effectively a memory leak because characters were getting deleted but their hats were being held onto by the table. If a player wore 7 accessories and died 100 times there would be 700 useless entries on the table because I forgot to clean it up when they died. The simplest solution is to make the keys weak so when the hats get deleted from the game the entry in the table gets deleted as well.
Very interesting but this method doesn’t work with Instances, so how did you make it work?

You could simply use the CharacterRemoving and CharacterAdded events to do this. It is better to do these types of things explicitly, similar to what Kampfkarren said.
I wanted to avoid this because I figured utilizing the garbage collector would be faster than Lua space code.
List of Wanderer keys
- Edit source
- View history
- Unfreeze Key - Can unfreeze any statue and bring it to life. it can also refreeze it into a statue. It can freeze random objects into stone as well, as seen when it turned a man's food into stone as he tried to eat it. When the magic of the key is reversed, it can unfreeze things that shouldn't be turned to stone and freeze things that are stone or a statue back to normal.
- Teeny-Tiny Key - Can make things super teeny tiny. When the magic of the key is reversed, the objects that it shrinks grows back to their normal size.
- Doubling Key - Can make duplicates of people, animals, and objects. When the magic is reversed, the duplicates disappear until the original being remains.
- Pepper Key - Can create a thundercloud that rains peppers.
- Wave Key - Can control waves, streams, rivers, and other watery places.
- Weather Key - Can control weather. When the magic of the key is reversed, the weather returns to normal.
- Body Switch Key - Can switch peoples' and even animals' bodies. When the key's magic is blasted at the two who were zapped by it in the first place, their bodies switch back.
- Color Key - Can drain the color out of anything it takes and inserts the color into clothes or anything else. When the magic of the key is reversed, the colored return to their rightful owners.
- Marshmallow Key - Can make marshmallows pop out from the end and make s'mores with the Chocolate Bar Key and the Graham Cracker Key. It's magic can also create an avalanche of marshmallows.
- Graham Cracker Key - Can make graham crackers pop out from the end and makes s'mores with the Marshmallow Key and the Chocolate Bar Key.
- Chocolate Bar Key - Can make chocolate bars shoot out from it's end and make s'mores with the Graham Cracker Key and the Marshmallow Key.
- Strength Key - Can give anyone the strength of a giant. When the magic shoots at the person again, they lose their super strength.
- Greek Myth Key - Can bring Greek mythology characters to life. When the magic of the key is reversed, it can send the characters and even the modern objects they hold back onto the urns.
- Crystal Glass Key - Can turn things into crystal glass. When the magic of the key is reversed, the crystallized-objects turn back to normal.
- Boomerang Key - Can be thrown like a boomerang and then bring other things to the thrower.
- Talk to Animals Key - Can turn animals' barks, chirps, yips, and growls into human words.
- Mega Mega Monster Key - Can grow anyone into the size of a giant monster. It also has a puzzle that needs to be solved in order for the magic to be reversed and shrink the person or animal back to it's rightful size.
- Slow Mo Key - Can slow anyone or anything down. When the magic of the key is reversed, everything reverts back it normal speed.
- Bubble Key - Can create bubbles from it's end. When the magic of the key is reversed, the bubbles pop.
- Ice Castle Key - Can create a giant ice castle. When the magic of the key is reversed, the ice castle disappears.
- Vacuum Key - Can suck up anything it picks up. When the magic of the key is reversed, it can shoot out anything it has sucked in.
- Tick Tock Key - Can take the travelers back in time to fix things.
- Quick Bloom Key - Can make any plant run, walk, jump, skip and garden like a human
- Follow the Leader Key -
- Riddle Key - Lead whomever has it will lead wanders to the hidden chamber of Magic Keys and gives riddles for assistance
- Bouncey Bounce Key - Can make anything bouncy. When the magic is reversed, the effect goes away
- Wind Key - Can create and control the wind.
- Shadow Puppet Key - Can bring shadows to life. When the magic is reversed the shadow turn to normal.
- Super Duper Snooper Key
- Key in the Stone - A legendary key trapped in stone similar to the Sword in the Stone from King Arthur legends which when pulled is said they will become the King or Queen of their own Kingdom
- Become an Animal Key - Can turn any person into an animal. When the magic is reversed the human turns back.
- Grow Up Key - Can make anything grow up or become young including humans. When the magic is reversed the target turns back to its original age.
- Dino Key - Can restore dinosaur fossils into real dinosaurs. When the magic is reversed, the dinosaurs turn back into fossils.
- Furball Key - Can cause fur to grow excessively. When the magic is reversed the fur turns back to normal
- Haunted House Key - Can bring Haunted House decertations to life. When the magic is reversed, the decorations turn back to normal.
- Invisibility Key - Can turn things invisible. When the magic is reversed, the invisibility is removed.
- Guitar Key - Resembles an acoustic guitar and it raise the sun and moon with a special song played correctly. When played incorrectly, the must can cause whomever listens to it, extremely dazed and causes the sun and moon to rise and fall out of control
- Fairy Tale Key - Can pull fairytale characters from their stories and into the real world. When the magic is reversed, the characters go back into the book
- Magnet Key - Can create a magnetic field and bring anything magnetic towards the key
- Wake-A-Troll Key - Can wake up giant, sleeping trolls
- Build a Bridge Key - Can build a bridge. When the magic is reversed the build disappears
- Honey Key - Can create honey. When the magic is reversed the honey disappears.
- Popcorn Cannon Key - Can create popcorn rapidly. When the magic is reversed, the popcorn disappears.
- Water Balloon Key - Can create water balloons. When the magic is reversed, the balloon disappears.
- Salt & Pepper Key - Can create a cloud which releases salt and pepper. When the magic is reversed it disappears.
- Burping Key - Can make the target burp.
- Fizzy Key - Can make the target frizzy.
- Cha Cha Key - Can make the target dance to the cha-cha. When the magic reverses, the target stops dancing.
- Bubble Gum Key - Can create bubblegum on any surface. When the magic reverses, the bubblegum disappears.
- Banana Peel Key - Can create a pile of bandanna peels. When the magic is reversed, the banana peels disappears.
- Trampoline Key - Can create a trampoline. When the magic is reversed, the trampoline disappears.
- Butterscotch Pudding Key - Creates a pile of butterscotch pudding on the ground. When the magic is reversed, it disappears.
- Island Maker Key - Can create whole islands. When the magic is reversed the island disappears.
- Ferris Wheel Key
- Whipped Cram Key
- Wrapping Key - Can wrap anything up into a present
- Flippety Key - Can make people could doing gymnastic
Gallery [ ]
")

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Philos Trans R Soc Lond B Biol Sci
- v.376(1817); 2021 Feb 1
Neuromodulation of the mind-wandering brain state: the interaction between neuromodulatory tone, sharp wave-ripples and spontaneous thought
Claire o'callaghan.
1 Brain and Mind Centre and School of Medical Sciences, Faculty of Medicine, University of Sydney, Sydney, Australia
2 Department of Psychiatry, University of Cambridge, Cambridge, UK
Ishan C. Walpola
James m. shine, associated data.
This article has no additional data.
Mind-wandering has become a captivating topic for cognitive neuroscientists. By now, it is reasonably well described in terms of its phenomenology and the large-scale neural networks that support it. However, we know very little about what neurobiological mechanisms trigger a mind-wandering episode and sustain the mind-wandering brain state. Here, we focus on the role of ascending neuromodulatory systems (i.e. acetylcholine, noradrenaline, serotonin and dopamine) in shaping mind-wandering. We advance the hypothesis that the hippocampal sharp wave-ripple (SWR) is a compelling candidate for a brain state that can trigger mind-wandering episodes. This hippocampal rhythm, which occurs spontaneously in quiescent behavioural states, is capable of propagating widespread activity in the default network and is functionally associated with recollective, associative, imagination and simulation processes. The occurrence of the SWR is heavily dependent on hippocampal neuromodulatory tone. We describe how the interplay of neuromodulators may promote the hippocampal SWR and trigger mind-wandering episodes. We then identify the global neuromodulatory signatures that shape the evolution of the mind-wandering brain state. Under our proposed framework, mind-wandering emerges due to the interplay between neuromodulatory systems that influence the transitions between brain states, which either facilitate, or impede, a wandering mind.
This article is part of the theme issue ‘Offline perception: voluntary and spontaneous perceptual experiences without matching external stimulation'.
1. Introduction
Mind-wandering is a mental state where thoughts arise spontaneously, relatively free from constraints and intentions [ 1 ]. Behaviour that is shaped by prior intentions, action plans and external constraints necessarily narrows the scope of possible states available to a system [ 2 , 3 ]. By contrast, mind-wandering suggests a widening of possibilities and a system untethered to constraints imposed by the external world. It reflects a system engaged in abstract, descriptive processes, shifted away from immediate sensorimotor goals or interactive behaviour with external affordances [ 4 , 5 ]. These characteristics are reflected in the phenomenology of mind-wandering: free-wheeling, undirected thoughts, with variable content and unpredictable trajectories. Such system properties and phenomenological characteristics are not unique to mind-wandering, but feature across the related phenomena of creativity, dreaming and hallucinations [ 6 – 8 ]. Collectively, these modes characterized by a lack of constraints on thoughts, and on the transitions between thoughts, are termed ‘spontaneous thought' [ 1 ].
As mind-wandering captured the attention of cognitive neuroscientists, the default network quickly became front and centre [ 9 – 11 ]. Although a more complex story has emerged, as an increased activity within the default network, and its relative engagement with attentional, control and sensorimotor networks, are the brain activation patterns most consistently associated with mind-wandering [ 1 , 12 – 14 ]. Yet, despite the reasonably detailed picture we now have of the attendant brain networks, we know very little about what might trigger mind-wandering and what neurobiological mechanisms control transitions in and out of the mind-wandering state.
Ongoing behaviour and cognitive function are strongly determined by fluctuations in brain state [ 15 ]. These states reflect an interaction between sensory input and intrinsically generated activity. In this way, brain state is dually determined by environmental engagement and by endogenously generated rhythms [ 16 ]. Despite this equilibrium process, most of our understanding of cognition has derived from studying brain activity and behaviours in response to external stimuli. Such ‘behaviour-yoked' paradigms tend to overlook the influence of underlying intrinsic brain states, which continuously shape and constrain ongoing cognitive, sensory and motor processes [ 17 ]. Indeed, many brain states, particularly during quiet periods of stillness or sleep, are dominated by stochastic, intrinsic activity patterns that are not driven by external inputs [ 18 , 19 ]. Endogenously generated activity seems particularly relevant to mind-wandering, which by its nature arises in a spontaneous, undirected manner, often with little or no identifiable influence from external stimuli. Here, we identify a hippocampal brain state, the sharp wave-ripple (SWR), as an intrinsically driven brain state that is particularly conducive to mind-wandering.
A key mechanism for regulating brain states are broadly projecting neuromodulatory systems [ 15 , 20 , 21 ]. The influence of ascending neuromodulatory systems plays a vital role across all aspects of behaviour, including higher order cognitive function [ 22 , 23 ]. By modulating the relatively fixed structural connectome of the brain to meet environmental challenges, neuromodulators support our capacity to flexibly transition across diverse modes of behaviour [ 24 , 25 ]. Chemical neuromodulation from these systems alters the properties of target cells by increasing, or decreasing, the likelihood of their firing (i.e. altering their gain; [ 26 ]). Neuromodulators can exert their influence over varying timescales, acting on target neurons to hyperpolarize or depolarize them, or to alter the plasticity of their synapses [ 27 ]. The resulting change in responsivity of target neurons is referred to as a change in neural gain, which can be thought of as a measure of neural signal amplification [ 28 , 29 ]. These properties of neuromodulators can be distinguished from the faster, direct signalling provided by classical neurotransmitters, such as GABA and glutamate [ 30 ].
While there are many families of neuromodulatory neurotransmitters, we focus on four of the major ascending neuromodulatory systems: acetylcholine, noradrenaline, serotonin and dopamine ( figure 1 ). Although each neuromodulatory system has unique characteristics, certain general principles apply. Despite their extensive projections, each system is capable of remarkably specialized regional effects; the systems are self-regulating, via actions at auto-receptors and descending projections; and, finally, they follow an inverted-U shaped curve (known as a Yerkes-Dodson-like function [ 31 ]), whereby efficacy of performance is governed by an optimal level of neuromodulatory influence, with too much or too little resulting in deleterious effects on behaviour [ 25 ].

Ascending neuromodulatory pathways. The ascending neuromodulatory systems and their projection nuclei, which send broad projections to different parts of the brain. ( a ) The cholinergic system, projections from the basal forebrain, including the septal nuclei (top) and basal nucleus of Meynert (bottom); and from the brainstem pedunculopontine/laterodorsal tegmental complex; ( b ) the noradrenergic system, from the locus coeruleus; ( c ) the serotonergic system, from the dorsal (top) and median (bottom) raphe nuclei; ( d ) the dopaminergic system, from the substantia nigra (top) and ventral tegmental area (bottom). (Online version in colour.)
To date, the limited attention that neuromodulators have received in mind-wandering has focused on the influence of tonic noradrenaline levels (e.g. see [ 32 , 33 ]). Extending these ideas, we examine the hypothesis that a balance between neuromodulatory neurotransmitters shapes brain states that promote spontaneous thought. We argue that tonic activity within specific combinations of neuromodulatory projection nuclei acts to influence the transitions between brain states that either facilitate (or interfere with) the ability to mind-wander. In addition, we describe how neuromodulatory tone might influence the phenomenology of spontaneous thought. We focus initially on the hippocampal SWR as a brain state that can trigger mind-wandering, highlighting the neuromodulatory influences that promote this hippocampal rhythm. We then describe more global neuromodulatory signatures that shape the subsequent evolution of the mind-wandering brain state.
2. Hippocampal sharp wave-ripples: a brain state for mind-wandering
(a) hippocampal sharp wave-ripples and their functional relevance for mind-wandering.
The link between behaviour and brain states has been well studied in the hippocampus, particularly with respect to theta and SWR hippocampal brain states, which both have their own characteristic oscillatory rhythms and behavioural correlates [ 16 , 19 ]. During awake, task-engaged behaviours, hippocampal activity is dominated by theta oscillations, promoting sensory processing and information sampling. By contrast, during slow-wave sleep and quiescent periods of wakefulness, and also during consummatory periods or following receipt of a reward, distinct neural patterns known as SWR complexes occur in the hippocampus ( figure 2 ). The sharp wave refers to high-amplitude depolarization of a large subset of CA1 neurons, driven by excitatory, recurrent activity in the upstream CA3 region; these sharp waves commonly co-occur with ‘ripples', which are brief, high-frequency oscillations in large populations of CA1 pyramidal neurons [ 18 , 34 ]. Importantly, SWR bursts are not induced but ‘released' in the absence of suppression mechanisms such as particular patterns of neuromodulatory tone [ 18 , 35 ]. The synchronous discharge of these pyramidal neurons in the hippocampal–entorhinal output pathway has powerful diffuse effects, leading to both up- and downregulation of widespread network activity [ 36 , 37 ]. Of particular relevance is that SWRs are consistently associated with hippocampal-prefrontal activation [ 38 ], and ripples have been linked to selectively increasing ongoing activity in the default network [ 39 ].

Hippocampal brain states. An example of the hippocampal theta rhythm and the sharp wave-ripple (SWR). Illustrated beside each recording is the associated behavioural state: locomotion and task engagement (theta), and a quiescent behavioural state, conducive to mind-wandering episodes (SWR). (Online version in colour.)
SWRs are a critical event in memory consolidation and retrieval. SWR neuronal activity can correspond to a previous experience, constituting a time-compressed ‘replay' or reactivation of that event. This activity may reflect events relevant to the immediate environment or can be drawn from more remote memories [ 40 ]. Coordinated hippocampal–cortical activity following SWRs is thought to promote ‘systems consolidation', whereby hippocampally mediated associations are progressively embedded within the synaptic weights that comprise neocortical networks, a process driven by the recurrent retrieval of memories over time [ 41 ]. This process is predicted by two-stage models of memory formation, where the encoding of new memories in the hippocampal formation occurs during active waking, and consolidation is promoted via random hippocampal reactivation during offline states of rest and sleep [ 42 ].
There is a good deal of flexibility in how these neuronal sequences are reactivated, suggesting that mere replay does not capture their diversity of function [ 43 ]. Part of the richness and diversity of SWR sequences may derive from the fact that they are often preceded by neocortical activity, consistent with the possibility that their content may be biased by information held in long-term memory [ 44 , 45 ]. During SWRs, neuronal sequences can be recapitulated in the forwards or reverse order of an experienced event [ 46 ], and activity can reflect novel combinations of past experiences or predicted future actions not previously experienced [ 47 – 49 ], sometimes comprising completely random, novel sequences [ 50 ]. Therefore, apart from a role in consolidating memories and retrieving them to aid immediate decision-making and planning, SWRs may also contribute to our ability to re-combine stored memories and simulate novel scenarios—abilities that underpin imagination [ 34 ]. The potential for SWRs to evoke memory and imagination and to activate the default network, raises the intriguing possibility that they may act as a trigger for episodes of mind-wandering.
(b) Hippocampal sharp wave-ripples drive default network activity and may promote conscious thought content
Hippocampal rhythms are capable of driving widespread neuronal activity. By synchronizing and coordinating large populations of hippocampal neurons, hippocampal waves can orchestrate the co-activation of distributed neuronal ensembles throughout the subcortex and neocortex [ 19 , 51 ]. This makes them ideally suited to support memory-related processes, which rely on information encoded across anatomically distinct brain regions [ 52 ]. Common neural regions are engaged by various memory-related processes, including remembering the past, simulating the future and imagination [ 53 ]. Regions activated during these processes reliably involve the default network and its context-dependent engagement with sensory, attentional and control networks [ 54 ]. Mind-wandering uses neural regions similar to those that support memory processes; however, it is initiated spontaneously and sustained in an undirected manner, which differs from deliberative, goal-directed processes that can typify memory retrieval and future planning [ 1 ]. The SWR hippocampal brain state is one such mechanism that can recruit neural regions involved in memory-related processes, but in a spontaneous, undirected way.
Dense hippocampal projections to core regions of the default network, including retrosplenial and posterior cingulate cortices, make this network a primary target for propagating hippocampal SWR activity [ 39 ]. Spontaneous SWR activation in the hippocampus and hippocampal–entorhinal output pathway has the potential to engage the default network more broadly, supporting the conscious elaboration of memories and simulations. One hypothesis is that the SWR acts as a subconscious search mechanism, which can bias subsequent thought content by priming the relevant circuits and assembly sequences necessary for an item to enter into consciousness [ 18 ]. Such biasing may occur via pattern completion processes, whereby a stored memory trace is activated with only a partial or degraded cue [ 55 – 58 ].
Pattern completion is classically attributed to the CA3 region of the hippocampus, on the basis of the recurrent collateral connections of its pyramidal cell population and its attractor dynamics [ 59 – 62 ]. This recurrent circuitry provides the basis of an ‘auto-associative network', capable of instantiating attractor dynamics that promote stable firing patterns. More concretely, the associativity of the network means that an entire neuronal population encoding an episodic memory may be activated based on activation of any subset of that population [ 63 , 64 ]. These dynamics support the highly associative nature of episodic memory and they are well described by two-stage models of memory recollection. Such models predict a priming of relational associations in the hippocampus—a rapid and unconscious process—which can either end, or evolve into a slower, conscious process associated with hippocampal–cortical interaction [ 65 , 66 ]. Pattern completion processes may also contribute to the itinerant and often highly associative nature of mind-wandering [ 67 ], where we can drift across a loosely connected train of thought, or certain contexts spontaneously evoke-related memories or imaginings. A memory recalled during pattern completion may trigger further pattern completions, which would support dynamic thought trajectories with variable content, but at the same time allow for thematic relationships and partial associations to persist across consecutive mental states [ 68 ]. Converging evidence from human studies shows that a considerable proportion of mind-wandering tends to be unconstrained or ‘freely moving' [ 69 ], and that when external demands are reduced, thoughts can tend towards novel, exploratory associations [ 70 , 71 ]. Hippocampal CA3 sharp wave activity has been linked to pattern completion attractor dynamics [ 72 ], providing a mechanistic link between the SWR brain state, and these associative mnemonic processes that may underpin mind-wandering.
It is currently unclear to what extent SWRs, which represent only a small fraction of hippocampally mediated activity in the awake state, are associated with conscious recollection versus a subconscious, mnemonic process [ 43 ]. Indeed, much of what we know about SWRs is based on rodents and non-human primates, so we lack detailed descriptions of their phenomenological content. Importantly, however, there is a good deal of human functional magnetic resonance imaging (fMRI) evidence demonstrating the spontaneous reactivation of previously learnt information during rest periods [ 73 ]. An even closer link with animal studies was recently established, with fMRI evidence for sequential offline replay identified in the human hippocampus following a non-spatial decision-making task [ 74 ]. Furthermore, decoded magnetoencephalography signals have been identified that are consistent with the temporally compressed replay of abstract rule sequences learnt during a task [ 75 ]. Importantly, these replay events coincided with ripple-band power increases (120–150 Hz) and could be localized to the medial temporal lobe, consistent with the hippocampal–neocortical regions activated during SWRs. Only very recently was the conscious content of hippocampal SWRs directly probed in human subjects, using intracranial electrophysiological recordings in neurosurgical patients. A transient increase in SWRs was observed just prior to the spontaneous free recall of images, which was related to SWR rates measured when participants originally viewed the images [ 76 ]. The anticipatory nature of their occurrence suggests a role for SWRs in the initiation of spontaneous recollection, providing evidence that SWR activation has the potential to influence the content of conscious thought.
A critical role for the hippocampus in influencing both the frequency and content of mind-wandering has been highlighted in recent work showing that hippocampal atrophy is associated with reduced mind-wandering in dementia [ 77 ] and reduced diversity in terms of content in individuals with selective bilateral hippocampal damage [ 78 ]. The SWR may be one mechanism that underpins the contribution of the hippocampus to mind-wandering. However, there is no reason to assume that all SWR events should provoke mind-wandering, as they are likely to be functionally heterogeneous, and both the anatomical locations of SWRs within the hippocampus [ 79 ] and timing of the sharp wave to ripple coupling [ 80 ] lead to different patterns of downstream brain activation. It could be anticipated that only certain subtypes of SWRs might promote mind-wandering.
Based on the evidence reviewed above, we suggest that SWRs represent a compelling candidate for a brain state that may trigger mind-wandering. The SWR hippocampal brain state meets many criteria relevant to mind-wandering: it is associated with relatively quiescent behavioural states; activity is spontaneously evoked and can be propagated via the default network; and it is functionally associated with recollective, associative, imagination and simulation processes. Through these mechanisms, SWRs may trigger episodes of mind-wandering, which are subsequently sustained and elaborated via the default network, and its engagement with attentional, control and sensorimotor networks [ 1 ]. In the next section, we summarize the relationship between SWRs and neuromodulatory tone, and the broader role of neuromodulators in larger scale brain dynamics, identifying how these might promote and sustain the mind-wandering brain state.
3. Neuromodulatory influences over hippocampal sharp wave-ripples and the global dynamics that sustain the mind-wandering brain state
The occurrence of SWRs is strongly determined by hippocampal neuromodulatory tone, suggesting that interactions between the neuromodulatory system and hippocampal SWRs may be a key component in the neuropharmacological signature of mind-wandering. Coupling between the hippocampus and subcortical neuromodulatory structures may provide an important means of orchestrating the brain-wide dynamics that follow SWRs [ 81 , 82 ], which are likely to be instrumental in sustaining mind-wandering. Neuromodulators are also important for constraining the brain-wide dynamics that shape the evolution of a specific mind-wandering episode. That is, although a specific set of circumstances may be required to trigger an SWR, the dynamic recruitment of different neuromodulatory systems may influence how a given mind-wandering episode plays out over time. Here, we describe the major neuromodulatory systems of the brain ( figure 1 ), and how they interact with the presence (or absence) of SWRs and the evolution of the mind-wandering brain state.
(a) Cholinergic system
Acetylcholine is released in widespread brain areas from projection nuclei in two main sites: the basal forebrain cholinergic system and the brainstem cholinergic system ( figure 1 a ) [ 83 – 86 ]. The basal forebrain cholinergic system includes cells in the medial septal nucleus and nucleus basalis of Meynert, which send projections to the neocortex and limbic system. Brainstem cholinergic neurons are clustered in the pedunculopontine tegmental nucleus and laterodorsal pontine tegmental nuclei, which together project to the thalamus, basal ganglia and basal forebrain.
The cholinergic system plays a major role in modulating brain states owing to its ability to promote high-conductance states in the cortex [ 87 ], which are associated with high-frequency cortical activation, reduced low-frequency fluctuations and less synchronized activity among neuronal populations [ 16 ]. The high-conductance state is linked to enhanced sensory processing, improved task performance and increased task engagement [ 88 – 92 ] and in the past has been described as desynchronized (though see [ 16 , 93 ]).
Cholinergic tone is at its highest during wakeful, task-engaged behaviours and also during rapid eye movement (REM) sleep, suggesting that the cortical activation in high cholinergic states need not necessarily be associated with behavioural activation [ 15 ]. Although the cholinergic system innervates many brain regions, there is increasing recognition that its projections are relatively localized [ 94 – 96 ], meaning that it influences target regions much more precisely than was previously thought [ 97 ]. In this way, while acetylcholine can have global effects that promote an activated brain state, it also plays highly specific roles in cognitive function. Acetylcholine promotes encoding, attentional selectivity and sensory processing, via various physiological effects that serve to improve the signal-to-noise ratio of neural processing and enhance processing of extrinsic/feed-forward inputs, relative to intrinsic feedback [ 86 , 98 ].
Cholinergic innervation of the hippocampus mainly derives from the septohippocampal pathway, with projections from the septal area (medial forebrain, rostral to the corpus callosum) innervating all regions of the hippocampus [ 99 , 100 ]. In the hippocampus, acetylcholine increases the amplitude of the theta rhythm, a state which is comparable to the high-conductance state in the cortex [ 16 ]. By contrast, the presence of acetylcholine is known to suppress SWRs, which has been demonstrated in vitro [ 101 ] and in both awake and anesthetized animals [ 102 ]. High cholinergic tone has been shown to simultaneously promote theta rhythms and suppress SWRs [ 72 , 102 ], suggesting that hippocampal theta functions in an antagonistic manner to SWRs as a function of cholinergic input. This is consistent with the opposing behavioural and cognitive correlates of the theta and SWR states. High cholinergic levels in the hippocampus, promoting the theta state, favour externally driven sensory processing and engagement with the environment [ 103 ], whereas the lower cholinergic tone promoting the SWR state supports internally driven processes [ 104 ]. The low cholinergic state enhances intrinsic hippocampal dynamics, allowing internal connections to be reorganized and strengthened based on previously encoded associations [ 105 ]. Taken together, a relatively low cholinergic tone in the hippocampus would bias intrinsic hippocampal dynamics over extrinsic input, promoting the spontaneous activation of SWRs.
At the global level, similar to the SWR hippocampal brain state, we hypothesize that mind-wandering should be associated with relatively low cholinergic tone. As described earlier, despite projecting to many areas of the cerebral cortex, cholinergic projections follow a highly topographic and differentiated, rather than diffuse, organization [ 106 ]. This organizational feature supports functionally and spatially selective signalling [ 107 ]. Cholinergic activity promotes selective neuronal population coding and a cortical network that is less driven by global fluctuations from diffuse inputs [ 96 ]. This is achieved by selectively boosting the neuronal gain in target regions, enhancing feed-forward connections and network specificity [ 108 – 110 ]. A direct prediction from these studies is that at the macroscopic network level, heightened cholinergic tone should promote segregated (i.e. tightly connected communities of brain regions with weak inter-connections) patterns of information processing [ 111 ]. Such segregated topology is optimized for functionally selective operations and sensory processing, which contrasts with the distributed, integrated information processing that would support mind-wandering.
A possible contention with the notion that relatively low cholinergic tone promotes mind-wandering is that cholinergic tone is also highest during REM sleep, and there are recognized phenomenological and cortical activation similarities between mind-wandering and dreaming [ 7 , 112 ]. However, contrary to the long-held view that dreaming is synonymous with REM sleep, dreaming is also known to occur during non-REM/slow-wave sleep [ 113 – 116 ]. While non-REM dreams often have a shorter and fragmented quality, they can also exhibit phenomenological characteristics indistinguishable from REM dreams, being longer, vivid and with a self-narrative [ 117 ]. When drawing parallels between dreams and mind-wandering, it is also important to note that neuromodulatory tone differs significantly across waking versus sleep. Serotonergic and noradrenergic systems considerably reduce their activity in non-REM and become inactive in REM, whereas cholinergic systems are virtually silent during non-REM, becoming highly active in REM [ 6 , 117 , 118 ]. Together, this highlights that similar phenomenological experiences can occur across waking and non-REM/REM brain states, each associated with very different neuromodulatory levels. In this respect, the low cholinergic state promoting SWRs may be considered one route to mind-wandering in the awake state, while different combinations of neuromodulators drive this phenomenology in other brain states.
(b) Noradrenergic system
The locus coeruleus (LC), situated deep in the pons, is the primary source of noradrenaline in the brain. This small nucleus has widespread projections innervating most brain regions, with the exception of the basal ganglia, and it provides descending input to the spinal cord and autonomic nuclei [ 119 ] ( figure 1 b ). Highly collateralized LC projections can release noradrenaline via non-synaptic or paracrine mechanisms, enabling diffuse influence over cortical activation and behavioural arousal [ 118 ]. At the larger scale brain network level, LC-noradrenaline activation facilitates reorganization of functional networks in response to changing environmental demands. Through the simultaneous action of noradrenaline at multiple target structures, ongoing functional interactions can be interrupted and reconfigured to promote a change in behaviour [ 120 ].
Early reports of LC-noradrenaline function focused on its role in general arousal and vigilance, based on its links with the sleep-wake cycle. However, increasingly more fine-grained functions have been related to the LC-noradrenaline system, including a central role in cognition and behavioural flexibility [ 121 ]. One way that noradrenaline regulates behaviour is by negotiating the balance between ongoing focus on a task and the need to shift focus to alternative options. Termed the ‘exploitation-exploration trade-off', this underpins an organism's ability to persist with a behaviour while it is useful and to explore more advantageous opportunities when use decreases. These modes of behaviour are mapped to temporally distinct firing patterns in the LC and changes in tonic noradrenaline levels [ 122 ]. When tonic noradrenaline levels are optimal, phasic firing occurs in response to task-relevant stimuli, and this is associated with task focus and good performance. Following a Yerkes-Dodson-like function, optimal task performance is typically associated with the middle range of tonic noradrenaline levels. Lower tonic levels are associated with low arousal and reduced alertness, which negatively impacts task performance. High tonic levels are associated with distractibility and an exploratory mode, facilitating the organism to disengage and pursue another behaviour.
Limited in vitro evidence using hippocampal slices suggests a receptor-dependant modulatory role of noradrenaline in SWRs, with α 1 adrenoreceptor activation associated with SWR suppression and SWR expression associated with β 1 adrenoreceptor activation [ 123 , 124 ]. Consistent with this, during certain SWR subtypes brain-wide activity shows downregulation of the LC, suggesting a modulatory role from this system in SWR events [ 80 ]. These studies raise the possibility that hippocampal noradrenaline levels may have both suppressive and potentiating effects on the occurrence of SWRs.
The LC densely innervates the hippocampus and regulates cellular excitability, cellular reorganization, synaptic plasticity and long-term potentiation, influencing all aspects of memory formation [ 125 – 128 ]. In addition to facilitating encoding, the LC-noradrenaline system is reactivated in the window after initial learning, at the stage of offline memory consolidation [ 129 , 130 ]. This phasic reactivation may relate to hippocampal SWR events. There is indirect evidence for this phenomenon, as delayed LC excitation after learning was found to occur exclusively during slow-wave sleep, a state in which SWRs are known to occur [ 130 ]. During slow-wave sleep, noradrenergic activity is at a relatively low tonic level; however, the LC is not completely silent [ 131 ]. The LC fires transient bursts during slow-wave sleep that coincide with hippocampal SWRs [ 132 , 133 ]. This suggests that phasic bursts of noradrenergic activity during states of relatively low (but not silent) noradrenergic activity may be conducive to SWR events in slow-wave sleep [ 134 ]. We speculate that similar noradrenergic factors—i.e. those that occur when one is awake but not particularly focused or stressed—may be conducive to SWR events in the awake state.
Co-activation of the noradrenergic system during replay could serve to promote plasticity and stability in the distributed cell assemblies reactivated during an SWR, contributing to systems consolidation in the offline state [ 121 , 130 , 135 ]. The noradrenergic release associated with an SWR-mediated phasic increase in LC activity would act to bring both the cortex and hippocampus into a more excitable, highly conductive state that would promote cross-regional interactions [ 136 , 137 ]. At the systems-level, the activation of relatively low-affinity α 1 adrenergic receptors promotes a ‘reset' of large-scale networks [ 120 ]. In this way, the noradrenergic activity coinciding with an SWR may be sufficient to reset cortical dynamics and in turn promote a brain state conducive to mind-wandering.
The effect of noradrenaline at the brain network level can be contrasted with the cholinergic system. The broad reach of gain modulation from the noradrenergic system is a key mechanism for enabling dynamic reorganization of cortical networks [ 138 ]. Noradrenergic function has been linked to increases in the strength and clustering of functional connectivity [ 29 ] and increased network integration [ 139 ], suggesting a role for noradrenaline in promoting distributed information processing across widespread brain regions, via its capacity for diffuse alterations in response gain [ 111 ]. Relatively higher global tonic noradrenaline, with respect to cholinergic tone, may promote increased integration across the brain to facilitate the coordinated information processing required to sustain episodes of mind-wandering.
(c) Serotonergic system
The serotonergic system innervates most brain regions, with the majority of ascending projections arising from the dorsal and median raphe nuclei in the brainstem ( figure 1 c ) [ 140 , 141 ]. Serotonergic projections arborize across diverse brain regions, with some providing paracrine transmission that enables diffuse influence over target regions. The serotonergic system acts on a large and diverse family of receptors [ 142 ], adding to the diverse functional capacity that is already conferred by its diffuse projections [ 143 ]. The highly selective distribution of receptor subtypes across cortical layers means that in addition to the ability to modulate brain-wide dynamics, the serotonergic system can exert very precise modulation over specific neuronal populations [ 144 ].
There is in vitro evidence that high levels of hippocampal serotonin can suppress SWRs [ 145 ]. In vivo , recordings from the serotonergic median raphe region (which projects to the entire hippocampal formation) showed that many of these neurons were inactive at the time of SWRs [ 146 ]. Furthermore, optogenetic excitation and inhibition of median raphe neurons respectively suppressed and enhanced SWR activity [ 146 ]. During certain SWR subtypes, brain-wide activity shows downregulation of the serotonergic dorsal raphe, in keeping with a modulatory role from this system in SWR events [ 80 ]. Therefore, inhibition of serotonergic hippocampally projecting neurons may be a critical factor in promoting the occurrence of SWRs. This action probably occurs via modulation of serotonergic 5HT 1A receptors, which are abundant in the hippocampal CA1 subregion [ 147 , 148 ] and are typically associated with inhibitory G-protein-coupled effects [ 149 ].
The concentration of serotonin in different brain regions could also influence the evolution of mind-wandering episodes after an SWR has occurred. There is compelling evidence that serotonin is involved in the alteration of global dynamics in a way that is consistent with the phenomenology of the mind-wandering brain state. Serotonergic psychedelics, such as lysergic acid diethylamide (LSD), psilocybin and ayahuasca/dimethyltryptamine (DMT), provide unique insights into the impact of brain serotonin upon the neurophenomenology of perception, as well as the boundaries between spontaneous thought and dreaming. While psychedelics do not act exclusively upon serotonin receptors, extensive work has revealed the dominant role for a specific serotonin receptor subclass (5HT 2A ) in psychedelic neurophenomenology [ 150 , 151 ]. Recent work has suggested that 5HT 2A agonists may shift the brain into a more entropic or anarchic mode of processing, in which the brain shifts between states in an irregular manner that is much less tethered to the external world than normal, waking consciousness [ 152 , 153 ]. By selectively increasing the gain of layer V pyramidal-tract-type cells (the major output population of the cerebral cortex), 5HT 2A agonists are suggested to flatten the attractor landscape [ 153 ], which means that novel patterns can form in the place of neuronal ensembles that are typically activated according to well-established firing patterns. This is consistent with the novel associations that may form during mind-wandering, raising the possibility that a mind-wandering episode occurring in the context of higher cortical serotonin may follow more novel, associative trajectories.
These lines of reasoning suggest dual effects of serotonin, with intermediate levels in the hippocampus promoting SWRs and higher levels in the cerebral neocortex promoting the evolution of a mind-wandering episode. Speculatively, these effects can be partly explained by actions at different serotonergic receptor subtypes (5HT 1A versus 5HT 2A ). This highlights the complexity of neuromodulatory influences over behaviour, as actions at different subtypes have varied and nuanced effects, which are mostly unexplored in relation to mind-wandering.
(d) Dopaminergic system
Dopamine may serve to promote SWRs in order to bias encoding and retrieval of salient or rewarding events [ 43 ]. There is a well-established role for dopamine in the formation and consolidation of memories, most notably via dopamine-driven mechanisms of hippocampal long-term potentiation. Midbrain dopaminergic neurons primarily modulate the hippocampus via a loop involving direct projections from the ventral tegmental area (VTA) to the hippocampus, which itself outputs (via the subiculum) excitatory projections to the nucleus accumbens, inhibiting the ventral pallidum and releasing VTA dopaminergic neurons from tonic inhibition [ 154 , 155 ]. In contrast with the diffusely branching collaterals of the noradrenergic and serotonergic systems that innervate multiple areas, dopaminergic projections (like cholinergic projections) are relatively more segregated and modulate more specific brain regions ( figure 1 d ) [ 156 ]. Consistent with this mechanism, the dopaminergic innervation of the hippocampus from the VTA is relatively sparse, potentially allowing it to have more specific, nuanced effects on hippocampal function [ 157 ]. Recently, an additional source of hippocampal dopamine was identified, as neurons projecting from the LC to the hippocampus co-release both noradrenaline and dopamine [ 158 , 159 ].
In relation to SWRs, direct application of dopamine to hippocampal slices in vitro results in a long-lasting increase in SWR frequency [ 160 ]. Furthermore, activity in the dopaminergic VTA-hippocampal pathway at the time of encoding enhances subsequent offline SWR activation [ 161 ]. This is consistent with earlier work showing enhanced SWR activity following rewarded outcomes [ 162 ]. Hippocampal dopamine at the time of experiencing novel, salient or rewarded events may therefore bias the content and frequency of subsequent SWR activity [ 163 ]. The role of dopamine may also extend to reinforcing reactivated sequences during offline periods, as dopaminergic input coordinates with hippocampal activity during SWRs in the offline state. This is demonstrated by reward responsive VTA neurons coordinating their firing with hippocampal SWR replayed sequences during offline quiet wakefulness [ 164 ].
These findings suggest a dual role for dopamine in increasing the likelihood of SWR reactivation of particular experiences, as well as further reinforcing reactivation patterns offline. While this presumably occurs as a means of promoting consolidation of salient events into long-term memory, it suggests that dopaminergic tone in the hippocampus may impact the subsequent spontaneous recall of information and, in this way, influence the content and the reinforcing aspects of mind-wandering. A link between the dopaminergic system and mind-wandering may substantiate recent theoretical assertions that a goal of mind-wandering is to generate potentially rewarding cognitive affordances [ 165 , 166 ].
(e) Interactions between neuromodulatory systems may trigger hippocampal sharp wave-ripples and sustain the mind-wandering brain state
Based on the evidence reviewed above, we propose that the combined influence of neuromodulators defines the likelihood that an SWR will occur in the hippocampus, and by extension, the propensity for an individual to enter into a mind-wandering episode ( figure 3 ). Lower cholinergic tone facilitates an overall dampening of sensory processing, disengagement from the external environment and a relatively quiescent behavioural state. This promotes hippocampal SWRs, which may trigger a mind-wandering episode. Similarly, low levels of hippocampal serotonin also facilitate the occurrence of SWRs. By contrast, high dopaminergic tone in the hippocampus can enhance the activity of SWRs and make it more likely for a neuronal sequence to be reactivated in the future, via reinforcing properties. Finally, phasic bursts during relatively low levels of noradrenaline may promote SWRs. Together, these patterns suggest that mind-wandering should occur during periods of relative quiescence (i.e. low cholinergic and serotonergic tone) but moderate arousal (i.e. low noradrenergic tone) in which a rewarding stimulus is chanced upon (i.e. heightened dopaminergic response), either via an external or internal cue ( figure 3 a ; teal sphere).

Neuromodulatory interplay that promotes the hippocampal sharp wave-ripple (SWR) brain state. It shows the balance of neuromodulatory tone that promotes hippocampal SWRs, which by extension is the neuromodulatory balance that may trigger a mind-wandering episode. ( a ) The teal sphere reflects the pro-SWR zone, that is, the balance of hippocampal neuromodulatory levels that is conducive to SWR occurrence; ( b ) an example of a precise state-space trajectory following an SWR; ( c ) if the SWR triggers increased noradrenaline, this will probably lead to a ‘network reset' [ 120 ]; ( d ) if the SWR recruits low cholinergic tone, this will facilitate further sensory isolation; ( e ) heightened serotonergic tone can trigger non-SWR-related mind-wandering. DA = dopamine; ACh = acetylcholine; 5-HT = serotonin; NA = noradrenaline. (Online version in colour.)
Once triggered, the occurrence of an SWR has the potential to drive brain networks into a precise, information-rich (i.e. low entropy) configuration ( figure 3 b ). Specifically, a hippocampal ripple re-activates distributed neuronal ensembles that were either related to the neuronal sequence that was embedded into the network during the original encoding event, or through the Hebbian processes that occur over the course of learning. In this way, a ripple event would act to re-energise a unique constellation of regions that were tangentially related to some aspect of the replay or simulation, opening up particular ways to engage with the current brain state over time (e.g. by re-exploring the same memory, or combining it in new ways).
As we have argued above, neuromodulatory tone plays an important role in shaping the likelihood of the hippocampus undergoing an SWR, and hence, how likely it would be to trigger a mind-wandering episode. However, the relative concentration of different neuromodulators within the pro-SWR zone will also probably play a role in defining the mind-wandering state. For instance, phasic noradrenergic activity at the time of the SWR may enhance the activation of distributed neuronal ensembles throughout the subcortex and neocortex, shaping the amount of integration present between otherwise disparate brain regions ( figure 3 c ). Higher concentrations of noradrenaline would probably facilitate a ‘reset' of the global brain state into one where a specific memory, association, or simulation has been primed into conscious contents, with the potential to trigger a mind-wandering episode. If the concentration of noradrenaline is too high (or low), this could render the phasic burst less (or more) impactful, as the phasic activity can potentially be obscured against the tonic background activity of the LC.
Once a mind-wandering episode has been triggered, how (or if) it evolves will also be further influenced by the neuromodulatory tone in the epoch immediately following a SWR. For instance, the continued low cholinergic tone in the basal forebrain system could downregulate the impact of incoming sensory input and bias the system towards intrinsic processes ( figure 3 d ). Lower cholinergic tone would also theoretically lessen the constraints on segregated network topology, allowing for more flexible integration across tangentially related neuronal ensembles [ 111 ]. Noradrenergic tone further determines the level of engagement with the current external or internal environment. If noradrenaline levels are optimal for whatever cognitive or behavioural task is at hand, it is likely that a person will maintain their focus on that task. However, if levels are still within an intermediate range but outside of task-optimal levels, then the triggered mind-wandering episode may take hold and allow the person to engage in spontaneous thought. Together, the cholinergic and noradrenergic systems have considerable influence upon overall brain states [ 167 ], working in cooperation to shape the extent to which we are driven by engagement with the external world, versus being driven by intrinsically generated activity.
While inhibition of hippocampal serotonin facilitates the occurrence of SWRs, wider-spread activation of the serotonergic system may shape the evolving content of spontaneous thought, which may not necessarily be triggered by an SWR ( figure 3 e ). Based on work using 5HT 2A receptor agonists [ 152 , 153 ], there is evidence that elevated serotonergic gain in layer V pyramidal-tract-type cells may release neuronal populations from their stereotypic firing patterns allowing novel patterns to form, consistent with the novel, unpredictable trajectory of certain spontaneous thought episodes. This mode of mind-wandering is consistent with a high-entropic state, as opposed to the low entropy state that may follow an SWR.
4. Relationship between neuromodulation and the phenomenology of spontaneous thought
Several lines of evidence link our proposed model of neuromodulatory influences with the phenomenology of spontaneous thought. We have suggested that low-to-intermediate levels of noradrenaline, albeit outside of task-optimal levels, are conducive to mind-wandering. In agreement with this, reduced pupil diameter (a proxy for tonic LC function [ 168 ]) indicative of low levels of arousal is associated with an increased frequency of hippocampal SWRs [ 88 , 169 ]. Evidence from human studies supports the idea that mind-wandering may occur when tonic noradrenaline levels are outside of task-optimal levels, as studies using pupillometry have identified off-task thought in the context of both elevated and reduced baseline pupils [ 32 , 33 , 170 – 173 ]. We suggest that spontaneous thought occurring in the context of relatively low tonic noradrenaline/low arousal most resembles the fleeting and transitory nature of mind-wandering. At the more extreme end of the low arousal/low tonic noradrenaline spectrum, thoughts may become increasingly disjointed and transient, with less awareness of thoughts, which at its extreme may be experienced as a mind-blanking state (i.e. thinking of ‘nothing') [ 174 – 176 ].
By contrast, relatively higher levels of tonic noradrenaline and arousal that facilitate an exploratory mode of behaviour may also engender an exploratory mode of spontaneous thought. This could be realized as exploratory creative thinking, which is subject to greater deliberative constraints than mind-wandering but still considered within the family of spontaneous thought [ 1 ]. In keeping with this, during divergent thinking—a creative thought process where many possible associations are explored—larger baseline pupil levels predict the generation of original ideas [ 177 ], suggesting a link between higher tonic noradrenaline levels and exploratory, creative thoughts. This contrasts with studies where lowered noradrenaline levels (via β-adrenergic antagonists) improved convergent thinking (i.e. zeroing in on a single creative response) [ 178 ], suggesting that noradrenaline may influence the extent to which creative thought is exploitative (convergent) versus explorative (divergent). Relatively high tonic noradrenaline, which would be characterized by high global gain and increased integration across brain regions [ 117 , 152 , 153 ], may support a brain state that promotes the dynamic default-executive network coupling that has been associated with divergent thinking [ 179 , 180 ].
Based on work using 5HT 2A receptor agonists, we have also suggested that serotonergic activity may facilitate the formation of novel patterns of neuronal ensembles as a mind-wandering episode evolves. Psilocybin is shown to enhance the activation of indirect semantic associations [ 181 ], which fits well with a recent proposal that certain aspects of mind-wandering may be driven by unconstrained semantic associations [ 67 ]. Furthermore, LSD was found to increase individuals' susceptibility to imagining themselves experiencing novel, creative scenarios [ 182 ]. Dreamlike states, with vivid visual imagery and cognitive bizarreness, induced by psychedelics [ 183 – 185 ] are also consistent with the possibility of novel pattern formation. Although the psychedelic state may be an intensified form of spontaneous thought, similar effects probably contribute to the formation of novel associations in the mind-wandering brain state.
5. Concluding remarks
Mind-wandering induces a shift from a pragmatic mode where an organism explores its environment, to a mode where mental states are explored. This process may represent a key evolutionary development, emerging as more complex systems interacted with more complex environments, where such offline processing enabled consolidation and refinement of learnt associations without being under the pressure of immediate goal-directed pursuits [ 186 ]. Such processing enables organization (and re-organization) of knowledge about the world into a high-dimensional space to support adaptive and flexible behaviour—an idea that has been conceptualized as a ‘cognitive map' [ 187 , 188 ]. The recurrent offline instantiation of memories and novel simulations that occurs with SWRs makes them an ideal candidate for maintaining and modulating these cognitive maps [ 189 , 190 ]. This possibility is supported by recent work showing that replay events in humans can constitute abstract rule sequences, suggesting they may function as a mechanism to generalize knowledge across experiences [ 75 ]. Considering a link between SWRs and mind-wandering, the possibility emerges that an adaptive function of mind-wandering may well be to augment these abstract knowledge structures we cultivate over our lifespan.
Here, we have defined spontaneous thought in terms of the possible brain states that might trigger and sustain it, focusing on the role of neuromodulatory systems in shaping the mind-wandering brain state. It is clear that each neuromodulatory system discussed plays a role in orchestrating the mind-wandering state. Indeed, the interplay between ascending arousal systems cannot be underestimated, as all of these systems contribute to waking consciousness and there are substantial interactions between them [ 191 ]. Continued understanding of this interplay remains important across all attempts to link behaviour and cognition with neuromodulatory systems. Likewise, although we have not focused on it here, a key principle of neuromodulatory systems that enables their exquisite flexibility is that they receive top-down regulation via the very regions that they are modulating [ 192 , 193 ]. Considering these reciprocal interactions also remains an important focus for reconciling neuromodulatory influences over brain-behaviour states. Finally, it is important to note that we have discussed brain states in somewhat absolute terms. It is now appreciated that brain states can at times be a complex mix of overlapping global and local sub-states [ 20 ], which combine to influence behaviour and cognition.
The field of cognitive neuroscience has made great strides in establishing the brain networks recruited during the mind-wandering brain state; however, we are only in the nascent stages of understanding how neuromodulators affect this state. Much of what we have presented here remains speculative; however, we hope that these early ideas may provoke the future work necessary to uncover the nuanced roles that neuromodulators undoubtedly play in mind-wandering.
Data accessibility
Authors' contributions.
C.O. was involved in conceptualization, writing original draft, review and editing the manuscript, and making figures. I.C.W. was involved in conceptualization, review and editing the manuscript. J.M.S. was involved in conceptualization; review and editing the manuscript and making figures
Competing interests
We declare we have no competing interests.
C.O. is supported by a Neil Hamilton Fairley Fellowship from the National Health and Medical Research Council (no. 1091310). J.M.S. is supported by a Robinson Fellowship from the University of Sydney and a project grant from the National Health and Medical Research Council (no. 1156536).
9 Essential Tips for Wandering Jew Plant Care

Sharing is caring!
Have you been looking for information on Wandering Jew Plant Care? Well, look no further. Call it luck or, pure coincidence that you have landed on this piece of information.
The article contains most if not all the things you need to know about taking care of this beautiful indoor vining plant . Be it propagation, watering, temperature and light requirements, pruning, repotting, or fertilizer application, it’s all in here.
Furthermore, I have touched on pests and diseases affecting this colorful, fast-growing indoor trailing succulent plant as well as common physiological problems associated with it.
Want to learn more about this plant?
Let’s begin.
Wandering Jew Plant Care: How to Grow and Care for Inch Plant ( Tradescantia zebrine )
1. propagate from stem cuttings.
Propagating a wandering Jew plant from stem cuttings is easy and quick. What’s more, is that you don’t need a special rooting medium or hormone for successful rooting. You just need to root the cuttings in water or soil.
Let’s start with rooting a Jew plant in water. First, cut at least 6-inch long cuttings from the healthy stems of the plant.
Then remove the bottom leaves from each stem and place the stems in a glass of water. Ensure that the bottom leaf node is submerged. In just a few days, tiny roots start to show, and in about 4 to 6 weeks, new growth appears.
At this stage, your cuttings are ready for transplanting. Use an all-purpose potting mix to grow your plants.
When it comes to rooting the Jew plant in soil, start by snipping several cuttings from the ends of healthy branches of your plant. Then using a clean and sharp blade, make a 45-degree cut just under the leaf node.
The next step is to fill an approximately six-inch hanging basket or pot with an all-purpose potting soil to about an inch below the top of the pot.
Afterward, make four holes approximately 2 inches deep around the edges of the pot and one more hole at the center.
Plant one cutting in each hole and add more soil as you press gently to hold the cuttings in position. Keep the soil evenly moist by regular watering and expect a new, full-leafy plant in a few weeks.
Pro Tip: Planting more than one stem in a single container makes the container appear fuller.
The plant thrives best in good-drained soil and with just an all-purpose potting mix, you are good to go!
It’s also fine to use regular potting soil provided that the soil doesn’t get soggy. You can improve soil drainage and aeration by adding some pumice or perlite.
In another case, your soil might seem to dry too quickly as opposed to holding moisture. Under these circumstances, mix in some vermiculite, peat moss or, coco coir to help the soil retain moisture.
3. Sunlight
This groundcover plant is hardy to zones 9 through 12 and it’s a nice houseplant.
Keep your indoor Jew plant, in a spot with bright but indirect light for example in an east or west-facing window. Your plant, therefore, will get plenty of natural light in the morning or evening, and bright indirect sun for the rest of the day.
This is important because when grown in too little light, the leaves’ color appears faded while under too much light, the leaves are prone to scorching.
On the other hand, an outdoor-grown plant requires a bright but partially shaded environment. This way the plant gets some shade during the hottest hours of the day.
This also applies to when you move your indoor plant outside for the summer.
4. Watering
Even though this plant is drought tolerant , it requires regular watering or, it won’t survive long spells of dryness or wetness. So, practice good watering habits and keep the soil moist but not soggy.
Using your finger, check whether the topsoil feels dry, or use a soil moisture gauge to check if the plant needs watering. If it does, water the plant thoroughly until water comes out through the drainage holes in the pot.
Then empty the sauce to avoid the development of diseases such as root rot.
5. Fertilizer Application
Although this plant doesn’t need to be fertilized, feeding it once in a while will benefit the plant by replacing the nutrients lost every time you water it.
Apart from this, fertilizing also encourages the plant to bloom. Imagine enjoying the beautiful view of purple, pink or white flowers even in winter!
Hence, feed the plant each month in spring through summer with a liquid fertilizer formulated for houseplants mixed at half strength.
Furthermore, this plant can be sensitive to chemical fertilizers so, use organic plant foods such as compost tea or a general-purpose fertilizer. Adding slow-release organic granules in the soil works well too.
You can also use liquid kelp and fish emulsion but only on outdoor-grown Jew plants as the two formulations can get somewhat stinky when used indoors.
6. Humidity & Temperature
Humidity is another key requirement for a healthy and good performing Jew plant. This plant loves lots of humidity.
Although maintaining good humidity levels indoors especially during winter can be challenging, it is possible to increase the levels using a humidifier .
Alternatively, fill a pebble tray with water and place the pot on the tray (don’t allow the pot to sit directly on water). This way, humidity around the plant increases as the water evaporates from the tray.
When it comes to temperature, the plant prefers warm temperatures of about 50-80 degrees. And when the temperature goes above 90 degrees, you need to water the plant more frequently and provide some shade to bring down the temperatures.
During winter, you need to be careful especially when the temperature is in a 45 degrees range. The plant will tolerate the condition for a short period but will die if the condition prolongs.
For this reason, device a way to help keep your plant’s ambient temperature above 50 degrees, for instance, a cold frame or a piece of woven cloth.
Pro Tip: When humidity levels are too low, the plants’ leaves start to turn brown and eventually die.
7. Repotting
This plant is fast-growing and repotting is quite necessary whenever the plant becomes crowded in its pot/container.
To repot, first select a container that’s approximately 50% wider than the existing one. Then, use some fresh potting mix around the sides of that container, remove the plant from the current container/pot and place it into the new one.
Next, remove or add the potting mix as needed to keep the plant into place and fill to about 2 inches below the containers’ rim.
Finally, press the potting mix lightly to hold the plant into place.
With time, there is a likelihood of your plant becoming leggy. When this happens, prune back the plant and pinch off the stem tips as well.
This way, the plant produces two new growths from right below the pinched areas and the plant becomes bushier and healthier.
Also, prune off the long tendrils if you like to have your plant thick and compact.
9. Look out for Pests and Diseases
Spider mites.
These are some of the most common pests in Jew plants and they cause damage by sucking sap from the plant. They like dry and warm areas.
Get rid of spider mites by misting, keeping the humidity high, or wash the plant with water to knock off the pests.
Plus, remove the infested plant parts/areas or use a systemic insecticide when the infestation is severe.
Aphids are another sap-sucking pests in wandering Jew plants. They are tiny, pear-shaped bugs that congregate along the stems in large masses.
Under severe infestation, the plant gets weak and eventually dies. Get rid of aphids by wiping the pests off the plant using a moist cloth (this works for low infestation), or spray the plant with an insecticidal soap or neem oil when the infestation is high.
This is the most common disease in wandering plants. It is caused by either over-watering or, your soil holding too much water. So, if the soil drains well but there is an occurrence of root rot, reduce the watering frequency.
And if the problem is poor drainage, devise ways of improving soil drainage such as adding some perlite or compost to the soil.
Common Problems in Wandering Jew Plant
Leggy plants.
Your plant might become leggy as a result of lack of light, especially during winter. You should hence keep your plant at a spot where it gets the right amount of sun.
However, if you don’t have such a spot in your house, try using grow lights to compliment the available natural light.
Brown leaves on inch plant
The leaves on your inch plant turn brown due to lack of enough moisture or humidity. Also, as the leaves age, they start dying out in the middle and this makes them appear brown. When this happens, prune the vines to refresh the plant.
Faded color on leaves
Faded or dull color on leaves is an indication of too much light, not enough light, or a bug infestation. Therefore, provide the appropriate growing conditions for your plant.
Now that you have read this article, I bet the question of wandering Jew plant care is a thing of the past.
But most importantly, you have seen the things that must be done correctly for a healthy plant.
For example; choosing an appropriate soil/potting mix, selecting the best propagation method, proper watering and, applying the correct fertilizer and at the right time.
Apart from this, pruning and, repotting crowded plants as well as protecting the plants from pests and diseases is a great way of ensuring you have a good–performing wandering plant.
Oh! And don’t forget to keep the environment humid enough.
What has been your experience growing wandering plants?
Share with us in the comments.
University of Wisconsin-Madison
Home Guides
1 thought on “9 Essential Tips for Wandering Jew Plant Care”
I ONCE READ THAT YOU SHOULD LET YOUR WATER SIT FOR 24 HOURS BEFORE USING IT TO WATER YOUR PLANT. ANY TRUTH TO THIS? ALSO, HOW DO YOU RECOMMEND ROTATING THE PLANT FOR EVEN SUNLIGHT. THANK-YOU, GG
Comments are closed.
Search code, repositories, users, issues, pull requests...
Provide feedback.
We read every piece of feedback, and take your input very seriously.
Saved searches
Use saved searches to filter your results more quickly.
To see all available qualifiers, see our documentation .
- Notifications
Releases: WanderingPhilosopher/Windows-KeySubtractor
Windowskeysubtractorv2.3.
Fixed a printing error.
WindowsKeySubtractorV2.2
New features added:
Completion percentage. -z flag; with -z you are telling the program to print only the x coordinate, not the 02 or 03 y coordinate parity.
To use on windows; download zip folder, unzip, run program in the same folder with the included .dll file.
Fixed count when -a was selected and added a percentage meter.
WindowsKeySubtractorV2
Version 2. Some added features; now you can either add or subtract ONLY from the public key, or add and subtract at the same time (as it was before.)
To use, download zip folder, unzip, and setup your batch file how you want it. NOTE: must keep libgmp-10.dll in same folder as the .exe file, unless you have libgmp-10.dll loaded on your PC and set in the Path/environments.
Windows Version 1.0
Download and unzip. File contains the .exe, a .dll, and an example batch file.
IMAGES
VIDEO
COMMENTS
List of All Weak Piece Codes. Below is a list of codes that will definitely help you in your journey across the seven seas. Make sure you enter the codes as they are because they can be case-sensitive. Moreover, redeem these codes as fast as you can. You never know when they will expire. Credit: XO Vanity. ACTIVE CODES.
Game - https://www.roblox.com/games/9487450292/UPDATE-5-Weak-PieceHey guys I hope you enjoyed the video/stream so make sure to hit the Subscirbe and Like but...
About Press Copyright Contact us Creators Advertise Developers Terms Privacy Policy & Safety How YouTube works Test new features NFL Sunday Ticket Press Copyright ...
The Weak Piece Trello contains details on updates, maps, NPCs, merchants, swords, fruits, fighting styles, items, boss spawns, and some general info on the game. It's a great way to get a lot of information at a glance. It is a bit simpler than a Wiki, because everything is on the same page and easily accessible!
In cryptography, a weak key is a key, which, used with a specific cipher, makes the cipher behave in some undesirable way.Weak keys usually represent a very small fraction of the overall keyspace, which usually means that, a cipher key made by random number generation is very unlikely to give rise to a security problem. Nevertheless, it is considered desirable for a cipher to have no weak keys.
;Sea warrior 10Minutesี;Shank 15Minutes;Kenshin 20Minutes;Ice Lord 25Minutesี;EnelGod 10Minutesv
The Key Monument was a gift to San Francisco from James Lick. One of the richest men in California, Lick died on October 1, 1876. In his trust, he set aside $60,000 for a monument to Francis Scott Key in Golden Gate Park to honor the composer of the Star Spangled Banner. It took over eight years before William Wetmore Story was awarded a ...
fight the 2 mini bosses in the back, collect the key item. You'll be forced to give up the key item regardless, so after the cut-scene... When you're near the entrance, turn around and go straight to the back of the cave and into the inner cave on the right you couldn't enter earlier. Watch the cut scene; Fight the 2 mini bosses again.
local coins = setmetatable({}, { __mode = "k" }) The idea is that the rest of your code is normal, but when the player leaves (and is parented to nil), it'll clear out the key. However, that's not the case because Weak Tables don't work with Instances. The actual solution is just to hook on PlayerRemoving and set the data to nil there.
your raw + your skill dame indicate your dealt damage while their defence (armour + star's ) show how much damage is passed on. a level 5 sword V level 1 sword is massively different in damage output. a max out Gray skill compared to a green skill, is different. combined your damage output is very pronounced.
Share your videos with friends, family, and the world
Visually, Wandering Sword makes good use of the Unreal engine to take 16-bit retro aesthetics and give them some extra muscle. Sprites move smoothly. Exploring through villages, caves, and other sites, as well as certain cutscenes, has just the right amount of parallax to it to give the feeling of classic JRPGs.
Follow the Leader Key -. Riddle Key - Lead whomever has it will lead wanders to the hidden chamber of Magic Keys and gives riddles for assistance. Bouncey Bounce Key - Can make anything bouncy. When the magic is reversed, the effect goes away. Wind Key - Can create and control the wind. Shadow Puppet Key - Can bring shadows to life.
List of all moves, lightness skills, and cultivation methods that I've found. Please comment about how to learn any moves I missed. Moves I have a manual for will a + next to them Sword Normal moves Basic Swordplay + Arching Swordplay - Consulted from Mu Haitang (Qingmu Sect Leader) Wudang Swordplay - Learned from Main Story or any Wudang sword user Mingjian Swordplay + Greypine Swordplay ...
Step 2: Trim weak or thin areas - Next, remove any thin, weak, or leggy sections of your wandering dude plant down to a lower leaf segment. You can either pinch them back with your fingers, or cut them using clean, sharp shears or snips. Cut back wandering jew just above a leaf joint.
1. Introduction. Mind-wandering is a mental state where thoughts arise spontaneously, relatively free from constraints and intentions [].Behaviour that is shaped by prior intentions, action plans and external constraints necessarily narrows the scope of possible states available to a system [2,3].By contrast, mind-wandering suggests a widening of possibilities and a system untethered to ...
basically just kill ace, he has 2 drops. One is the White orb which is for bisento, and The weapon drop is Excalibur. so just kill him as many times u can ti...
1. Propagate from stem cuttings. Propagating a wandering Jew plant from stem cuttings is easy and quick. What's more, is that you don't need a special rooting medium or hormone for successful rooting. You just need to root the cuttings in water or soil. Let's start with rooting a Jew plant in water. First, cut at least 6-inch long ...
To use on windows; download zip folder, unzip, run program in the same folder with the included .dll file. Fixed count when -a was selected and added a percentage meter. Version 2. Some added features; now you can either add or subtract ONLY from the public key, or add and subtract at the same time (as it was before.)
Weak Piece Codes (2022) | How to Redeem Codes in Roblox Weak Piece
A mutant species of Kung Fu Dugong that rove alone, away from the herd. They focus too much on hand-to-hand combat, and are weak to ranged weapons. Also, their serious nature makes them more likely to fall for jokes and surprise attacks.
📌Map https://www.roblox.com/games/9487450292/UPDATE1-Weak-Pieceเพลงต้นคลิป https://www.youtube.com/watch?v=L8ppv7i7Iwg 🟠ช่อง ...